Tuesday, May 17th 2011
Sons and daughters and differential parental investment
One of my favorite rhetorical tricks is asking my students a question that has an obvious answer based on cultural expectations, but is wrong. So every year, when I start to teach my students about parental investment, I ask:
Who is harder to raise, sons or daughters?
I’ve asked by a show of hands and with iClickers, over the years, and the room of 750 is almost unanimous: daughters are harder to raise. So, then I get off the stage and walk around a bit. What do you mean by that? I ask.
Girls cause more gray hairs.
Girls cause more trouble when they start to like boys.
Girls are more work, and cost more money, since they shop all the time.
Girls talk back more.
And of course, there is always the saying that girls steal some of their mother’s beauty.
So then I show them this:
 |
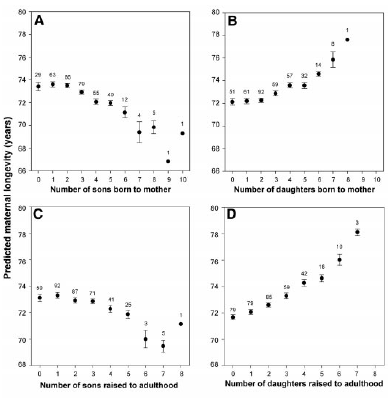
| From Helle et al 2002. |
Here is a graph of maternal longevity based on the number of sons or daughters they have. This data was based on a historical population from Finland from 1640-1870 using church records (Helle et al 2002). As you can see, the more sons mothers bear, the shorter their lifespans. You see the opposite for daughters. So sons have a negative impact, and daughters have a positive impact. This same trend has been found in records from a Flemish village (van de Putte et al 2003, 2004), where sons negatively impact lifespan but not daughters. Interestingly, data from church records from the field site where I work in rural Poland provides a slightly different picture: every offspring of either sex reduced lifespan by about 95 weeks (Jasienska et al 2006).
 |
| From Jasienska et al 2006. |
Once students see these graphs, they quickly realize what is going on. Generally speaking, girls help mothers more at home in terms of chores and alloparenting. And in many cultures, particularly the historical ones studied so far, sons are costly because parents invest more in them, to help launch their own families. Daughters, not so much. In the Polish population, there may be other factors where daughter investment is important, or it is just costly to have so many offspring and you have maternal depletion regardless of daughter help.
Sex bias in parental investment is an important part of understanding both the biology and culture of parenting, and the developmental trajectories of children. The Trivers-Willard hypothesis, which has been tested many times in humans and animals, suggests that parents should invest more in sons when conditions are good, and more in daughters when conditions are bad. That is, when you have lots of resource you should put it towards a son in order to increase the chances he will have high reproductive success, since his is assumed to be more variable and high effort could lead to high reward. But in periods of low resource, daughters are a good bet because they are more likely to have at least some reproductive success no matter what.
 |
| From Hrdy 1990. |
Of course, differential investment based on resources is further conflated in humans due to culture and, I would contend, our almost universal favor for patriarchy (Smuts 1995). Here is an image of an Indian family waiting at a clinic. There is a mother, an older son, and a twin boy and girl. Notice the extreme difference in health between the infant boy and girl – they are twins, yet the infant girl is emaciated. This is because in this population sons are always fed and cared for first, and whatever is left over, if there’s anything, is given to daughters.
So, parental investment can have real effects on the parent in terms of lifespan, and perhaps also their own future reproductive capabilities. Further, the conditions under which you may have children can vary, but how much a parent chooses to invest in their children varies too.
The piece of this that may be toughest to parse out, particularly in humans, is how the condition of the mother (or parents) can vary, and how that variation impacts the sex ratio of their children. In some species, like red deer, it is easier to imagine a mechanism: these animals have diapause, a period where their embryos are dormant until it is a good time to gestate and bear them. It is easier to insert some kind of selection process into a period where several embryos are all “frozen” and sex has been determined. But what about humans that produce singletons and invest huge, overlapping amounts of support to their children over decades? How would a sex bias based on maternal condition operate? And is there anything the offspring can do about it?
Changes in maternal breast size during pregnancy
It turns out that measurements as easy as stepping on a scale, and knowing your bra size, can begin to unpack the answer. First, a confession: I consider the author of this paper Andrzej Galbarczyk more than a colleague, but a friend. Andrzej is the graduate student who oversaw my Polish field site last season (Mogielica Human Ecology Study Site, director Dr. Grazyna Jasienska). He has translated consent forms and surveys for me and we’ve had many valuable and important conversations about my fieldwork. He is a smart, kind and thoughtful person and scholar. So, I let him see an early draft of this post to make sure I understood his point of view.
Galbarczyk performed an internet survey in Poland with 120 women, where he asked them to report their pre-pregnancy weight and bra size, their bra size directly after giving birth, and the sex of their offspring. He found two notable differences in these women: mothers of daughters weighed less before pregnancy, but had a greater changes in breast size during pregnancy.
The evidence about maternal pre-pregnancy weight is consistent with the Trivers-Willard hypothesis, as mothers who had sons were more likely to be heavier, and thus have more resource to invest. The second significant difference, that mothers with daughters had larger breasts after pregnancy, seems could be argued either way: Galbarczyk argues that it supports Trivers-Willard because mothers of sons could have been devoting more resource to growing their offspring rather than their breasts.
In other animals and primates particularly, mothers of male infants produce more energy-dense milk, yet mothers of female infants may produce a greater quantity of milk (Hinde 2009). And breast size is a pretty noisy signal of milk quality or quantity. So, what is the meaning of this difference in breast size?
Adaptation or physiological inevitability?
Galbarczyk suggests the difference is related to the evolutionary underpinnings of human female breasts. Women develop breasts around puberty, and though they certainly change in size and shape over time, keep them their whole lives. Other animals develop their mammary glands only shortly before lactating and then they regress again. Many contend that human breasts are an honest signal of fertility. This is at least partially confirmed by the correlation between breast size and estradiol concentrations (Jasienska et al 2006).
Galbarczyk thinks that the larger breasts seen in postpartum mothers to daughters may be a way to attract a mate for parental care. Perhaps this would help where she has given birth to the less-favored sex and needs to really convince him to participate; this could be a signal from the mother or the female fetus. Or maybe by appearing more attractive, she can have another reproductive opportunity, which would give her a chance to have a son.
You all know how I feel about evolutionary storytelling. In certain ways I do find this particular argument compelling, from the perspective of the Trivers-Willard hypothesis. But the evidence for the adaptive scenario around breast size is circumstantial.
Also, I don’t want this story to detract from some very interesting data: remember that Galbarczyk found that in this population, mothers of daughters weigh less before pregnancy, and develop larger breasts afterwards. Very cool. So perhaps we should consider a mechanistic, rather than adaptive explanation?
I have two thoughts about this, both related to androgens (androgens are the class of hormone that testosterone falls under). First, I wonder if there is an effect of fetal androgens from a male fetus on breast size. If so, mothers of daughters would have larger breasts simply because they aren’t having their breast tissue growth or density suppressed by androgens. It could simply be physiology that doesn’t have adaptive meaning.
Second, the mothers of sons were heavier before pregnancy. Heavier individuals tend to have higher circulating insulin levels, and the ovary can respond to higher insulin by producing more androgens (Poretsky 1991, Dimitrakakis et al 2004). So you could have a suppressive effect on breast size from that avenue as well. You don’t need an adaptive scenario for either of these mechanisms, just a consequence of how hormones work.
I would love to see Galbarczyk or someone else follow up on these thought-provoking results by measuring women, rather than relying on self-report, and by measuring their estradiol, progesterone and androgens. Understanding the different factors and motivations that lead to sex differential investment and outcome is a great field of study, and this work gets us thinking in a new direction.
References
Dimitrakakis C, Jones RA, Liu A, & Bondy CA (2004). Breast cancer incidence in postmenopausal women using testosterone in addition to usual hormone therapy. Menopause (New York, N.Y.), 11 (5), 531-5 PMID: 15356405
Galbarczyk A (2011). Unexpected changes in maternal breast size during pregnancy in relation to infant sex: An evolutionary interpretation. American journal of human biology : the official journal of the Human Biology Council PMID: 21544894
Helle, S. (2002). Sons Reduced Maternal Longevity in Preindustrial Humans Science, 296 (5570), 1085-1085 DOI: 10.1126/science.1070106
Hinde K (2009). Richer milk for sons but more milk for daughters: Sex-biased investment during lactation varies with maternal life history in rhesus macaques. American journal of human biology : the official journal of the Human Biology Council, 21 (4), 512-9 PMID: 19384860
Hrdy, S. (1990). Sex bias in nature and in history: A late 1980s reexamination of the “biological origins” argument American Journal of Physical Anthropology, 33 (S11), 25-37 DOI: 10.1002/ajpa.1330330504
Jasienska G, Nenko I, & Jasienski M (2006). Daughters increase longevity of fathers, but daughters and sons equally reduce longevity of mothers. American journal of human biology : the official journal of the Human Biology Council, 18 (3), 422-5 PMID: 16634019
Poretsky L, Seto-Young D, Shrestha A, Dhillon S, Mirjany M, Liu HC, Yih MC, & Rosenwaks Z (2001). Phosphatidyl-inositol-3 kinase-independent insulin action pathway(s) in the human ovary. The Journal of clinical endocrinology and metabolism, 86 (7), 3115-9 PMID: 11443175
Wednesday, April 6th 2011
When I was younger, periods were not a fun time, and I was plagued with dysmenorrhea, which is a fancy term for really bad cramps. In high school, I would often take 1000 mg of ibuprofen every four hours to alleviate symptoms to get through all my classes, band, sports practice, and homework (what, it took you this long to realize I was, and am, a dork?).
After having my daughter in 2008, and the thirteen months of lactational amenorrhea that followed it (lactational amenorrhea means absence of periods due to lactation), my periods resumed. Pain during my periods has almost totally ceased, but I have noticed more cycle-related variation in emotion. In particular, my patience and tolerance for rude behavior, and my tendency to cry sentimentally at even the lamest greeting card, skyrocket in my premenstrual phase. I already have low tolerance for rudeness, and I already cry easily. But something about progesterone decline — which is a normal process towards the end of ovulatory cycles — seems to make it harder for me to repress these behaviors in order to fit in culturally with those around me.
I tell this to you to say, I don’t doubt that hormones, and hormonal variation through the cycle, plays some role in variation in female behavior and emotion. And I find this kind of work inherently interesting. I hate to repeat myself, but you will find echoes of my structural and methodological concerns with evolutionary psychology in this post as well.
* * *
Durante et al (2011) observe that women spend more money on their appearance than men, and claim that this sex difference is cross-culturally consistent (I wonder, is this consistent across cultures without money?). In order to understand this sex difference, they wish to see whether spending or shopping behavior is dependent on cycle phase. Therefore the authors hypothesize that women choose sexier clothing during ovulation — “even if the women themselves are not consciously aware of this biological fact” (Durante et al 2011: 922), a problematic turn of phase if I ever saw one, but I’ll get to that later. They also consider the effects of priming a shopping woman with images of attractive women and hypothesize there is a greater effect of this priming on high-fertility women.
Methods
The participants were female undergraduates and were compensated with course credit or money. The authors claim the participants had no idea the study had anything to do with the menstrual cycle, but the participants had to use LH strips at midcycle to see when she was ovulating (this is a urine test to check for a luteinizing hormone peak, which comes before ovulation).
Here’s the important part, for me:
“The first urine test was scheduled 2 days before the expected day of ovulation. If an LH surge was not detected, women came back each day until an LH surge was detected or six tests had been completed, whichever came first” (Durante et al 2011: 924).
Here are my questions: what is 2 days before the expected day of ovulation? The follicular phase — that’s from menstruation to ovulation — is the most variable phase of the menstrual cycle (Fehring et al. 2006; Lenton et al. 1984). I wonder how many ovulations they missed because of this. Perhaps even worse, how many participants had six LH tests and didn’t have a detectable LH surge? It sounds like they were included in the project. But, they either ovulated before the authors started testing, or they had an anovulatory cycle. That means the authors were including participants in their study that weren’t ovulating… in a study of behavior during ovulation.
Participants viewed a made-up shopping website on a high-fertility (near the LH surge) and low-fertility (about eight days later) day, where they had to select ten items they would like to buy that day. They were randomized into two groups: one shown a site featuring casual clothes, the other featuring clothes and accessories. The clothing on these made-up sites were “pretested to be sexy” (Durante et al 2011: 925). While that is a phrase I never expected to write on this blog, the separate validation they did to determine sexy versus nonsexy clothing seems fine.
Results
Hypothesis 1: Near ovulation, women should be more likely to choose sexier and revealing clothing and other fashion items rather than items that are less revealing and sexy (Durante et al 2011: 923).
Women chose a greater percentage of sexy clothing and accessory items near ovulation: 59.8% ± 21.6 during ovulation, 51.3% ± 22.4 during low fertility. This was a statistically significant difference, but they did a repeated measures ANOVA, and I don’t understand why they didn’t do a paired t-test. Further, statistically significant or not, I question how meaningful it is when the averages are so close and the standard deviations almost completely overlap.
H2: Ovulation should lead women to be especially likely to choose sexier products when women are primed to compare themselves to attractive female rivals (Durante et al 2011: 924).
H3: There should be no differences in product choice between ovulating and nonovulating women when women are primed with unattractive women or men (Durante et al 2011: 924).
Follow-up studies primed sub-sets of participants (so a different cohort, same recruitment methods as above) to think about 1) attractive local women, 2) unattractive local women, 3) attractive local men, 4) unattractive local men. They did this by showing photographs of people who they claimed to be local and asking participants to rate their attractiveness.
When primed with attractive women, the percentages of sexy items chosen were 62.7% for ovulating women and 38.2% for low fertility women (I could not find standard deviations for these values so have no idea how much the two groups overlap). Priming with unattractive women, attractive men, or unattractive men produced no significant difference between low and high fertility women.
H4: Ovulation should lead women to choose sexier products when primed to think about local attractive women who constitute potential direct rivals. However, ovulation should not influence product choice when women are primed to think about women from distant locations because such women do not constitute direct rivals (Durante et al 2011: 924).
The authors used a different method for assessing fertility this time; they asked women their normal menstrual cycle length and counted back from menses to estimate when ovulation would be. So AGAIN, we don’t know how many women actually ovulated in this study, and we don’t know whether a significant portion of women were then grouped in the high-fertility group who shouldn’t have been.
This study is like the previous one in terms of photo priming, but this time the photos were said to be local or distant, and were of women only (so the four groups were local attractive, local unattractive, distant attractive, distant unattractive).
The authors claimed that the relationship between fertility, photo attractiveness and location was “marginally significant,” but the p-value was 0.09. That is, in fact, not significant, as significance is generally only considered under 0.05 unless you cheat and say your study is special and should consider a different limit (they don’t say this in their study).
That said, the only significant effect found of photo priming on high versus low fertility women was in the local attractive women group: high fertility women chose 65.8% sexy items versus low fertility’s 39.1% (I could not find standard deviations for these values so have no idea how much the two groups overlap). These results are almost identical to those found when priming women with attractive women without saying if they are local or not.
How biological are we talking here?
The authors claim a biological cause for the differences found above. And maybe there is, to some extent. But there are two major issues with the authors’ conclusions.
First, there is the major methodological flaw of including women who probably aren’t ovulating in their high-fertility group. Heads up to people who don’t study female physiology: women, even healthy women with “normal” cycle lengths, don’t ovulate every cycle. So if understanding a behavior during ovulation is important to your hypotheses, you need daily hormones on top of that LH test. Then, you know, if you can’t document ovulation, you need to exclude those women from your sample. Oh, and while we’re discussing methods, the authors don’t mention whether the participants were in a relationship or not, or what their sexualities were, or their races or socioeconomic statuses. These are all important to understanding variation in female-female competition (Campbell 2004). And since ornamentation is likely related to honest signals of health, it would be good to know waist to hip ratios, or BMI, or facial symmetry (Streeter and McBurney 2003) (hello, I’m handing someone a dissertation here! Just remember to cite me correctly).
But the second issue relates to the theme I saw throughout this paper, that changes in mood or choice behavior due to ovulation or presence of attractive women is a “biological fact.” Female-female competition is certainly found within human behavior, and behavior changes through the menstrual cycle. But is it fair to call these behaviors strictly biological, or should we have a more nuanced understanding of the interaction between biology and culture?
There are alternative cultural theories out there. Objectification theory proposes that there are consequences to living in a culture that sexually objectifies women: when women are continually appraised based on their looks, it leads to a disconnect between their body and individual (Moffitt and Szymanski 2011). This disempowers women and leads to them feeling as though their bodies exist for the pleasure of others. And if this is what women learn they have to offer others, and they seek affirmation, praise or attention from those around them, it makes sense for women to compete around attractiveness, particularly sexiness.
I would posit that shopping, particularly when primed with the image of an attractive woman, is a kind of objectification. So really, what Durante et al (2011) are measuring are the results of objectifying their study participants. Under these circumstances, a woman is more likely to start treating herself as an object to be evaluated on the basis of her appearance, so it makes sense that she would choose sexier clothing, in an effort to produce a culturally-appropriate, attractive body.
As the study stands, there is no way to parse out the impact of biology or culture — and many cultures encourage objectification, female-female competition and female attractiveness towards men. As for how that interacts with high versus low fertility samples… that’s the interesting part of this paper. If we can trust how the women are parsed. Which we can’t, since some of the high-fertility sample might not have been ovulating.
These high heels are made of deer antlers
The authors also seemed enamored with the idea of comparing their female participants to male animals. Twice they mention the idea that they want to determine whether sexy clothing is analogous to a peacock’s tail, a deer’s antlers, or a lion’s mane (really). These three examples, according to the authors, reflect a courtship function, a same-sex competition function, or both functions respectively. The authors go on to say that their results suggest that sexy dressing in women is like deer’s antlers, or, a same-sex competition function.
First, since when are a deer’s antlers only a same-sex competition function? Second, doesn’t it say something that they couldn’t find any examples of this kind of display in a female animal? This begs the question of why female humans do so much more displaying and maintenance of their appearance compared to other female animals, and again, this suggests interactions between biology and culture (Smuts 1995).
We can spin all the stories we want to explain why many human females make efforts to be physically attractive. And I do think Durante et al (2011) are on to something here as, despite methodological concerns they did find differences in high- and low-fertility choices. But if we continue to do this research on undergraduates in western contexts without sufficient hormone analysis, I’m unsure that its meaning extends beyond the participant pool.
References
Campbell, A. (2004). Female competition: Causes, constraints, content, and contexts Journal of Sex Research, 41 (1), 16-26 DOI: 10.1080/00224490409552210
Durante, KM, Griskevicius, V, Hill, SE, Perilloux, C, & Li, NP (2011). Ovulation, female competition, and product choice: hormonal influences on consumer behavior Journal of Consumer Research, 37 (6), 921-934
Fehring, R., Schneider, M., & Raviele, K. (2006). Variability in the Phases of the Menstrual Cycle Journal of Obstetric, Gynecologic, Neonatal Nursing, 35 (3), 376-384 DOI: 10.1111/j.1552-6909.2006.00051.x
Lenton EA, Landgren BM, Sexton L, & Harper R (1984). Normal variation in the length of the follicular phase of the menstrual cycle: effect of chronological age. British journal of obstetrics and gynaecology, 91 (7), 681-4 PMID: 6743609
Moffitt, L., & Szymanski, D. (2010). Experiencing Sexually Objectifying Environments: A Qualitative Study The Counseling Psychologist, 39 (1), 67-106 DOI: 10.1177/0011000010364551
Smuts, B. (1995). The evolutionary origins of patriarchy Human Nature, 6 (1), 1-32 DOI: 10.1007/BF02734133
Streeter, S. (2003). Waist–hip ratio and attractiveness New evidence and a critique of “a critical test” Evolution and Human Behavior, 24 (2), 88-98 DOI: 10.1016/S1090-5138(02)00121-6